Why do capable, well-intentioned teams so often fall into frustration, competition, or subtle conflict?
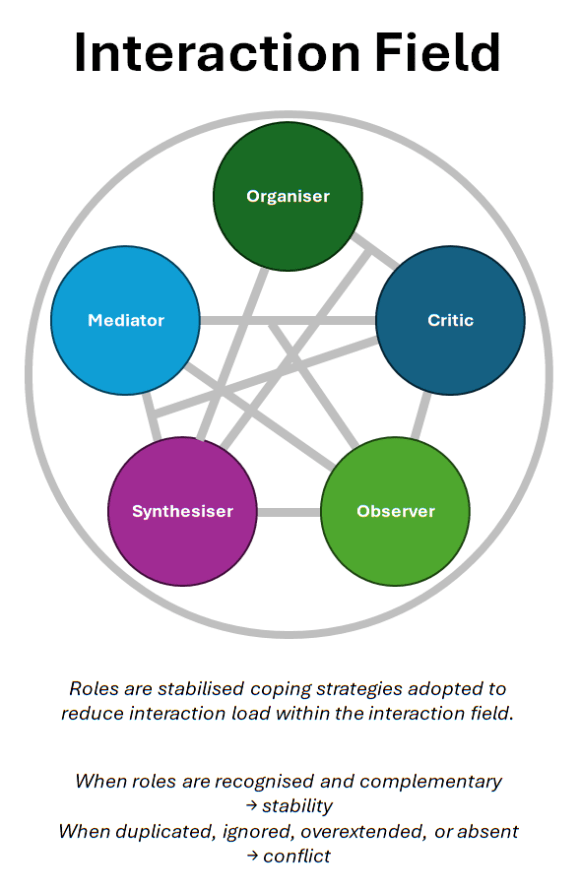
This article proposes a different way of understanding what is happening beneath the surface of group behaviour. It suggests that what we commonly call “roles” are not job titles or personality traits, but stabilised coping strategies that people adopt in order to manage the rising complexity of interacting within a group.
When these roles are recognised and complementary, teams work smoothly. When they are duplicated, ignored, overextended, or absent, tension and dysfunction appear, often without anyone understanding why.
The article also shows how the same pattern repeats across levels of systems: what appears as roles in teams appears as departments in organisations and as specialisations in economies and nations.
This provides not only a practical explanation for many common forms of team conflict, but a systems perspective on how cooperation can be improved within and between groups at every scale.
We often speak of “information” as though it floats freely in cyberspace or the human mind, detached from anything physical. Yet every bit of information, from the letters on this page to the thoughts in your head, is carried by matter or energy. This simple observation lies at the heart of cognitive physicalism, the view that cognition, communication, and social coordination are all thermodynamic processes.
Information Is Order
In physical terms, information is negative entropy; order among components of a system. When the atoms of a crystal, the base pairs of DNA, or the neurons of a brain are arranged in regular patterns, they hold information by reducing randomness. This definition, first clarified by Léon Brillouin and Erwin Schrödinger, gives information the same physical dimensions as entropy:
Energy provides the capacity for work (); information provides the form that directs that work. Together they make organisation possible.
How Physics Becomes Mind
In purely physical systems, energy and entropy simply flow. With life, informational structures emerge that regulate those flows. A cell maintains order by channelling chemical energy through genetic and enzymatic constraints. With evolution, feedback control grows more elaborate: nervous systems model the world, predict outcomes, and choose among options. Agency, the ability to act purposefully, appears when informational form controls energetic process.
At higher levels, the same principle produces cognition, language, and society. Neural firing, conversation, and economic exchange are all manifestations of energy flows organised by information.
Why Equations Matter
When information theory borrowed from thermodynamics, it kept Boltzmann’s equation but quietly normalised away the constant Doing so made information appear dimensionless; handy for communication engineers, but misleading for science. As Rolf Landauer later reminded us, information is physical: erasing a single bit requires energy and generates heat. Ignoring this fact masks the cost of learning, computing, and communicating; costs that become crucial when we extend systems thinking to living and social domains.
The Structure of Agency
Agency can be described in three physical layers:
Level
Description
Dimensions
Agentic information structure
pattern that directs energy
Agentic potential
information-structured energy capacity
Actualised agency
directed energy flow through time
Energy provides the means, information the form, and their coupling the act. Whether in a cell, a mind, or a society, the same dimensional hierarchy holds.
The Sun and the Spectrum of Agency
All terrestrial agency begins with the Sun. Photons striking chlorophyll are converted into chemical potential, which sustains metabolism, cognition, and eventually culture. Every thought, conversation, or social reform is therefore a distant echo of solar radiation; a transformation of sunlight into structured work.
The Cost of Thought and Change
Learning, decision, and communication are thermodynamic operations. Brain imaging shows energy consumption rising during problem-solving; each new memory reduces neural entropy while producing waste heat. The same principle scales up: cultural and institutional change require energy to reorganise shared information. Schools, media, and political movements are energetic engines for lowering societal entropy. When their energy supply falters, coherence and collective agency decline.
Why This Matters for Systems Science
Re-embedding information and agency in physics brings fresh clarity to systems thinking. It explains why order must be sustained by flows, why “effort” feels costly, and why every form of coordination, from metabolism to governance, depends on continual energy input. It also offers a bridge between natural and social sciences: the same thermodynamic grammar governs both.
As Ilya Prigogine showed, local order can grow even while global entropy rises. Life, mind, and society are all such dissipative structures, islands of organisation maintained by throughputs of energy and information. Understanding this continuity reminds us that progress itself carries an energetic price.
From Theory to Application
Recognising the physical nature of information could reshape how we approach education, technology, and governance. Policies and systems that ignore their energetic base risk collapse; those that respect it can harness energy more efficiently to sustain informational order.
Energy is the means, information the form, and agency the dance between them. Seen thermodynamically, every act of understanding is a small victory over entropy; a local flowering of order in the great energetic flow from the Sun.
Two Motivational Reflexivity Community groups have been created on LinkedIn and Facebook, where you can ask questions, share your experiences, provide feedback, and stay updated on developments. These groups are open to all who are interested in deepening their understanding of Motivational Reflexivity and connecting with others practicing this approach.
These resources and community spaces are free to access. Feel free to download, engage, and share the links with anyone who might benefit. Your participation and feedback are invaluable as we build a supportive community around this practice.
In the longer term, I am planning to produce guidelines for trainers, a dedicated website, and online training courses, all of which will be free to share and use. Their availability will be announced here and in the Facebook and LinkedIn groups.
The paper presents a comprehensive hypothesis that seeks to explain the nature of reality and how humans understand it, integrating foundational concepts from critical realism, systems theory, and causality. The hypothesis holds that reality can be viewed as a fractal-like structure, generated by underlying organising principles that operate at various ranks in a hierarchy. Starting from acausal foundational principles, the paper explores how systems interact, transfer matter, energy, and information, and contribute to the complexity observed at different levels of organisation. The hypothesis extends to the idea that human understanding is structured by organising principles that differ from reality’s, leading to distinct layers of comprehension reflected in scientific disciplines. The paper suggests that integrating these principles may help bridge gaps between disciplines, such as the disconnect between social sciences and the biological sciences. This unification has the potential to deepen our understanding of both the natural world and human social behaviour, while identifying new pathways for societal change.
In this part, the work of the English philosopher of science, Roy Bhaskar (1944 – 2014), and the English sociologist, Margaret Archer (1943 – 2023), is described and commented upon. Bhaskar’s contribution to the theory of society was twofold. Firstly, his “transcendental realism” dealt with the nature of science in general, and secondly, his “critical naturalism” with the social sciences in particular. The two terms were later conflated by his followers into “critical realism”, the philosophy of science of which he is now regarded as the founder. His transcendental realism is consistent with the author’s “Systems Theory from a Cognitive and Physicalist Perspective”. The latter was derived independently, largely from work on symbolic logic. However, Bhaskar also provides further insights that will be described in the paper. His work can be regarded as falling within the discipline of systems science, although Bhaskar makes little reference to systems. Regarding Bhaskar’s critical naturalism, I generally agree with this. However, there are details on which we diverge that will also be described. Archer’s main contributions to the theory of society were her explanations of social morphogenesis and reflexivity, both of which are also described and commented upon.